- O nás
- Naše aktivity
- Nabízíme
- Děkujeme

Partnerské weby
![]()
![]()
Obecná biologie lýkožrouta smrkového
Vloženo: 22.12.2020, Typ: Jiné, Vydáno: 22.12.2020
Kůrovci tvoří podčeleď Scolytinae spadající do čeledi Curculionidae (nosatcovití) z řádu Coleoptera (brouci). Kůrovců existuje zhruba 6000 druhů [1]. Většina z nich kolonizuje pouze mrtvé stromy, napomáhá jejich rozkladu a urychluje koloběh živin. Lýkožrout smrkový (Ips typographus) však při nízkých početnostech napadá stresované a umírající stromy (například po velkých bouřkách následovaných suchým a horkým létem) a ve vyšších hustotách si troufne dokonce i na strom zdravý. Kromě nejznámějšího lýkožrouta smrkového můžeme v českých lesích najít i lýkožrouta severského (Ips duplicatus), lýkožrouta menšího (Ips amitinus) či lýkožrouta lesklého (Pityogenes chalcographus).
.jpg)
Obr. 1: Samice lýkožrouta smrkového. Foto: Gilles San Martin, CC BY-SA 2.0
Lýkožrout smrkový se považuje za nejvýznamnějšího škůdce evropských smrkových lesů. První velká zdokumentovaná kalamita tohoto škůdce se odehrála na území České republiky, konkrétně na Šumavě, po velkých bouřích v letech 1868 a 1870 [2]. Odhaduje se, že mezi roky 1850 a 2000 bylo 8.1 % veškeré úmrtnosti stromů způsobené kůrovci, zejména pak lýkožroutem smrkovým[2]. Aktuální masová kalamita v řadě evropských zemí však nemá v historii obdoby. Těžba kůrovcových stromů často přesahuje až 50 % z celkového ročního objemu sklizně [3].

Obr. 2: Galerie vytvořené lýkožroutem smrkovým. Kůrovcem napadený strom poznáme podle přítomnosti závrtů na kmeni (ve výšce cca 1,3 metru a výše): těsně po náletu můžeme ještě pozorovat oranžovo-hnědé drtinky slepené pryskyřicí v okolí závrtu či na zemi u paty kmene. Jakmile zaprší, drtinky se spláchnou.
Dospělci lýkožrouta smrkového začnou být aktivní na jaře. Doba prvního jarního rojení závisí na teplotních podmínkách a pohybuje se v rozmezí od konce dubna po začátek června (ve vysokohorských oblastech). Samci se prokoušou pod kůru stromů, vytvoří snubní komůrku a svými feromony přitahují samičky. Jeden samec svými feromony přitahuje od jedné až po čtyři samice [4]. Po spáření samice budují dlouhé vertikální chodby, podél kterých kladou samostatně do malých prohlubní vajíčka [5]. Délka chodby a počet nakladených vajíček klesá s rostoucím počtem samic obývajících strom [6]. Při napadení stromu větším množstvím jedinců se samice mohou vynořit a opět zavrtat do stejného či jiného stromu a bez dalšího páření naklást tzv. sesterské pokolení [7]. Po vylíhnutí si larvy kolmo na matčinou chodbu (vertikálně) vytvoří krmící tunely na jejichž konci je kukelná kolébka, ve které se zakuklí (lýkožrout tedy patří mezi holometabola, tj. hmyz s proměnou dokonalou). U čerstvě vylíhlých dospělců dochází první ke sklerotizaci kutikuly (vytvrdnutí vnější schránky) u čehož se krmí lýkem stromu, poté k opuštění rodného stromu a kolonizaci stromu nového [8]. Průměrná doba vývoje od vajíčka po dospělce trvá 29 dní. Minimální teplota potřebná pro dokončení vývoje je cca 10,5 °C, maximální cca 40 °C. V teplotním rozmezí 15 °C až 25 °C se rychlost vývoje s rostoucí teplotou lineárně zvyšovala [9]. V České republice může být lýkožrout v závislosti na podmínkách univoltinní (s jednou generací za rok) či multivolinní (s více generacemi za rok, v ČR maximálně čtyři generace). Při více generacích za rok se stává, že někteří jedinci nestihnou dokončit vývoj a jsou nuceni přečkat zimu ve stádiu larvy či kukly a umírají (letální teplotní hranice pro larvy a kukly jsou mezi -13 C˚ a -17 ˚C) [10]. Počet generací lýkožrouta za rok výrazně ovlivňují faktory jako např. zeměpisná šířka, teplota či dostupnost potravy a očekává se, že budou nadále narůstat v reakci na klimatickou změnu [3].
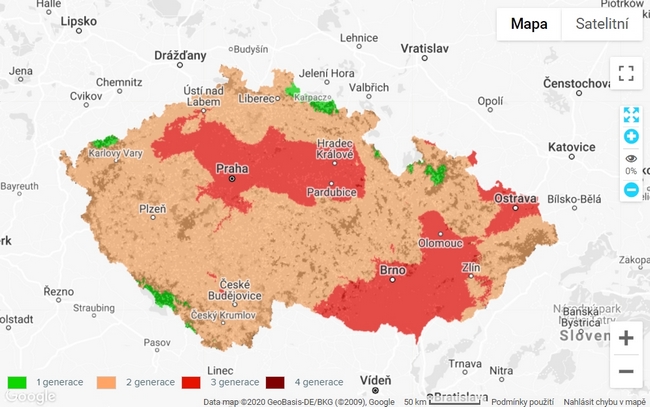
Obr. 1: Výskyt různého počtu generací lýkožrouta smrkového (Ips typographus) v České republice. Online zdroj: https://www.klimatickazmena.cz/cs/?l=64 [cit. 13. 10. 2020, 20:10]. metodika zde: https://www.klimatickazmena.cz/cs/metodika/model-phenips/
Diapauza se běžně vyskytuje u ektotermních živočichů jako strategie pro přečkání nepříznivých podmínek. Dochází při ní k pozastavení vývoje a snížení rychlosti metabolismu. U lýkožrouta může být diapauza obligátní či fakultativní. Brouci s obligátní diapauzou do ní vstupují bez ohledu na vnější podmínky a jsou považováni za striktně univoltinní. Lýkožrouti s diapauzou fakultativní do ní vstupují na základě změny délky fotoperiody a tím jsou potencionálně multivolinní [8]. Lýkožrout smrkový přečkává zimu jako dospělec buďto v kůře napadeného stromu nebo v opadu. Nedávné experimenty ukazují, že podíl populace brouků, kteří tráví diapauzu v kůře stromů se zvyšuje se snižující se nadmořskou výškou. Jedinci z jižních zeměpisných šířek a nižších nadmořských výšek nejsou během zimy nuceni opouštět kůru. Pro severní Evropu a vyšší nadmořské výšky jsou typické dlouhodobě nízké teploty vzduchu, které mohou způsobit mrazivé zranění [11].
[1] Reid M. L. (2004). Bark beetles. Encyclopedia of Forest Sciences. 119-126. doi:10.1016/B0-12-145160-7/00030-2
[2] Schelhaas, M.-J., Nabuurs, G.-J., & Schuck, A. (2003). Natural disturbances in the European forests in the 19th and 20th centuries. Global Change Biology, 9(11), 1620–1633. doi:10.1046/j.1365-2486.2003.00684.x
[3] Hlásny, T., Krokene, P., Liebhold, A., Montagné-Huck, C., Müller, J., Qin, H., Raffa, K., Schelhaas, M-J., Seidl, R., Svoboda, M., Viiri, H. (2019). Living with bark beetles: impacts, outlook and management options. From Science to Policy 8. European Forest Institute. 52 pp.
[4] Jeger M. et al. (2017). Pest categorisation of Ips typographus. EFSA Journal. Volume 15, Issue 7, Article number e04881. doi: 10.2903/j.efsa.2017.4881
[5] Doležal, P., & Sehnal, F. (2007). Effects of photoperiod and temperature on the development and diapause of the bark beetle Ips typographus. Journal of Applied Entomology, 131(3), 165–173. doi:10.1111/j.1439-0418.2006.01123.x
[6] Anderbrant, O. (1990). Gallery construction and oviposition of the bark beetle Ips typographus (Coleoptera: Scolytidae) at different breeding densities. Ecological Entomology, 15(1), 1–8. doi:10.1111/j.1365-2311.1990.tb00777.x
[7] Wermelinger, B., & Seifert, M. (1999). Temperature-dependent reproduction of the spruce bark beetle Ips typographus, and analysis of the potential population growth. Ecological Entomology, 24(1), 103–110. doi:10.1046/j.1365-2311.1999.00175.x
[8] Schebeck, M., Hansen, E. M., Schopf, A., Ragland, G. J., Stauffer, C., & Bentz, B. J. (2017). Diapause and overwintering of two spruce bark beetle species. Physiological Entomology, 42(3), 200–210. doi:10.1111/phen.12200
[9] Wermelinger, B., & Seifert, M. (1998). Analysis of the temperature dependent development of the spruce bark beetle Ips typographus (L.) (Col., Scolytidae). Journal of Applied Entomology Volume 122, Issue 4, June 1998, Pages 185-191. doi: 10.1111/j.1439-0418.1998.tb01482.x
[10] Kasumović L., Lindelöw A., Hrašovec B. (2018). Influence of predator abundance and winter mortality on reproduction of bivoltine populations of Ips typographus L. (Coleoptera: Curculionidae). Izvorni znanstveni članci – Original scientific papers. Šumarski list, 9–10 (2018): 473–479. doi:10.31298/sl.142.9-10.2
[11] Kasumović, L., Lindelöw, A., & Hrašovec, B. (2019). Overwintering strategy of Ips typographus L. (Coleoptera, Curculionidae, Scolytinae) in Croatian spruce forests on lowest elevation. Šumarski List, 143(1-2), 19–24. doi:10.31298/sl.143.1-2.2